Overview
- Three late australopithecine species — A. garhi, A. sediba, and A. deyiremeda — demonstrate that the period from 3.4 to 1.98 million years ago saw a diverse radiation of hominins, not a single linear chain of ancestors.
- Australopithecus garhi is associated with some of the earliest evidence of stone-tool-assisted butchery at 2.5 million years ago, while A. sediba displays a striking mosaic of australopith and Homo-like traits in its brain, pelvis, and hand.
- Australopithecus deyiremeda demonstrates that multiple hominin species coexisted in the same region of Ethiopia 3.4 million years ago, overturning the long-held assumption that A. afarensis was the sole hominin of its era.
For decades, the story ran as a tidy progression: A. afarensis begat A. africanus, which begat early Homo. Discoveries since the mid-1990s have thoroughly complicated this picture. Three species described between 1999 and 2015—Australopithecus garhi, A. sediba, and A. deyiremeda—reveal that the later australopithecine period was characterized by remarkable diversity, with multiple species occupying overlapping habitats and exhibiting different mixes of primitive and derived features.1, 2, 3 Together, they illuminate the bushy nature of the hominin family tree during the critical interval when Homo first emerged.
Australopithecus garhi: discovery and context
Australopithecus garhi was described in 1999 by Asfaw, White, and colleagues from a partial cranium (BOU-VP-12/130) recovered at the Bouri Formation in Ethiopia's Middle Awash.1 The name garhi means "surprise" in Afar.4 Found by Yohannes Haile-Selassie in 1997, the holotype dates to approximately 2.5 million years ago, a period then poorly represented in the hominin record.1

The Bouri Formation lies within the Middle Awash, an extraordinarily productive region that has yielded hominin fossils spanning more than five million years.5 The Hata Member preserves lakeside and river-margin sediments—shallow waters, floodplains, and gallery forests—a mosaic landscape supporting diverse resources.6
Anatomy and tool use of A. garhi
The A. garhi cranium combines features that distinguish it from all other known australopithecines. Cranial capacity (~450 cc) falls at the upper end of the australopithecine range.1 Most distinctive is the dentition: extraordinarily large premolars and molars exceeding A. afarensis, yet without the heavy facial buttressing and sagittal cresting of Paranthropus.1, 4 This combination of megadont teeth without robust cranial architecture suggested a lineage distinct from both the robust clade and the lineage leading to Homo.1
A nearby partial skeleton (BOU-VP-12/1), not definitively attributed to A. garhi, preserves forearm and thigh elements. If it belongs to the same species, it suggests limb proportions in which the forearm remained long while the femur had already elongated—a combination not seen in any other known hominin, implying lower limb elongation preceded forearm shortening.1
Perhaps most significant is the association with stone-tool-assisted butchery. Mammalian bones from the same level bear unmistakable cut marks and percussion marks indicative of defleshing and marrow extraction.6 Although no tools were found in direct association at Bouri, Oldowan artifacts of similar age come from nearby Gona.7 The proximity makes A. garhi a candidate for the earliest hominin engaged in tool-assisted meat processing, though other species may have been present.1, 6
Australopithecus sediba: a boy's discovery
The story of Australopithecus sediba begins with one of paleoanthropology's most charming anecdotes. On 15 August 2008, nine-year-old Matthew Berger stumbled upon a fossil clavicle while exploring around the Malapa cave site in South Africa's Cradle of Humankind.2 His father, paleoanthropologist Lee Berger, recognized it as hominin and initiated excavations that yielded two remarkably complete partial skeletons: MH1 (juvenile male, holotype) and MH2 (adult female).2 Announced in Science in 2010, the name sediba means "wellspring" in Sotho, reflecting Berger's hypothesis that this species might represent the wellspring of Homo.2
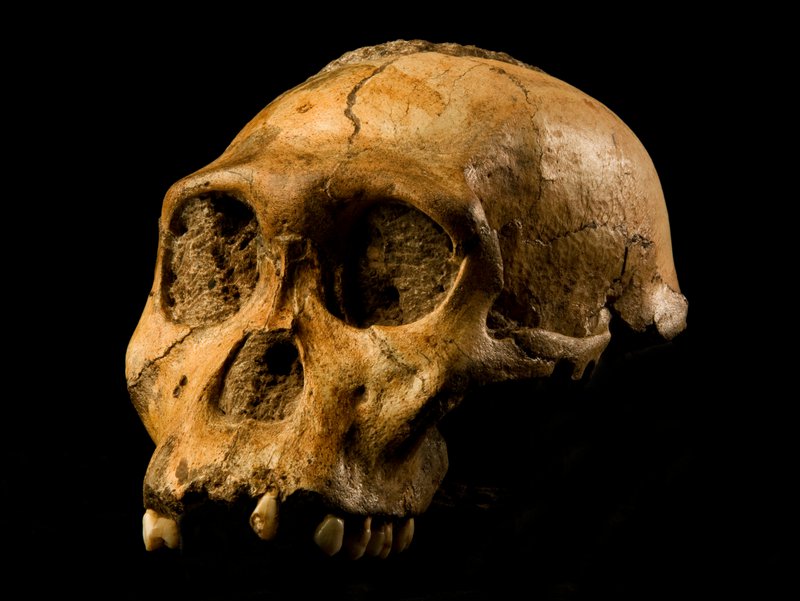
The two individuals appear to have fallen into a natural death trap—a deep shaft in the dolomitic cave—and were rapidly buried, resulting in exceptional preservation.2 Uranium-lead and paleomagnetic dating place the specimens at ~1.977 million years ago.8 This date falls after the earliest known Homo fossils, fueling debate over whether A. sediba could truly be ancestral to Homo or is instead a late-surviving australopithecine side branch.9
The mosaic anatomy of A. sediba
What makes A. sediba exceptional is the degree to which it combines australopithecine and Homo-like features within a single body. A 2011 series of studies in Science documented this mosaic across the brain, pelvis, hand, and foot.10, 11, 3, 12
The brain of MH1 had a cranial capacity of only ~420 cc—firmly australopithecine—yet the frontal lobes showed subtle reorganization, aligning more closely with Homo than with other australopithecines in principal component analysis.10 This suggested neural reorganization may have preceded the dramatic brain size increase of Homo, with profound implications for understanding how human cognition evolved.10
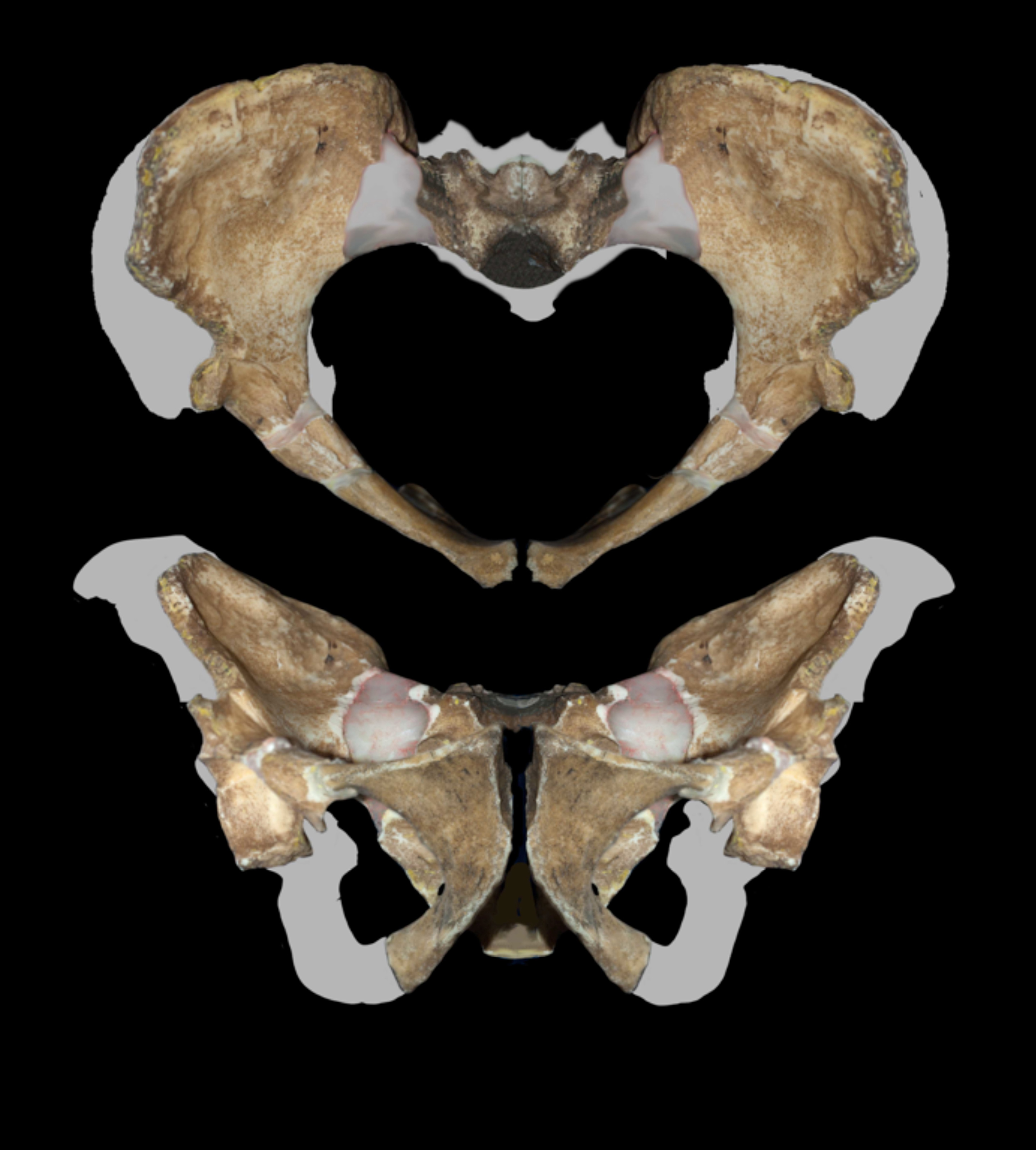
The pelvis, reconstructed from fragments of both MH1 and MH2, exhibited derived features suggestive of more human-like bipedalism than earlier australopithecines—even while retaining long arms suited for climbing.11 These pelvic changes occurred before brain expansion, challenging the hypothesis that the human-like pelvis evolved primarily to accommodate larger-brained infants.11
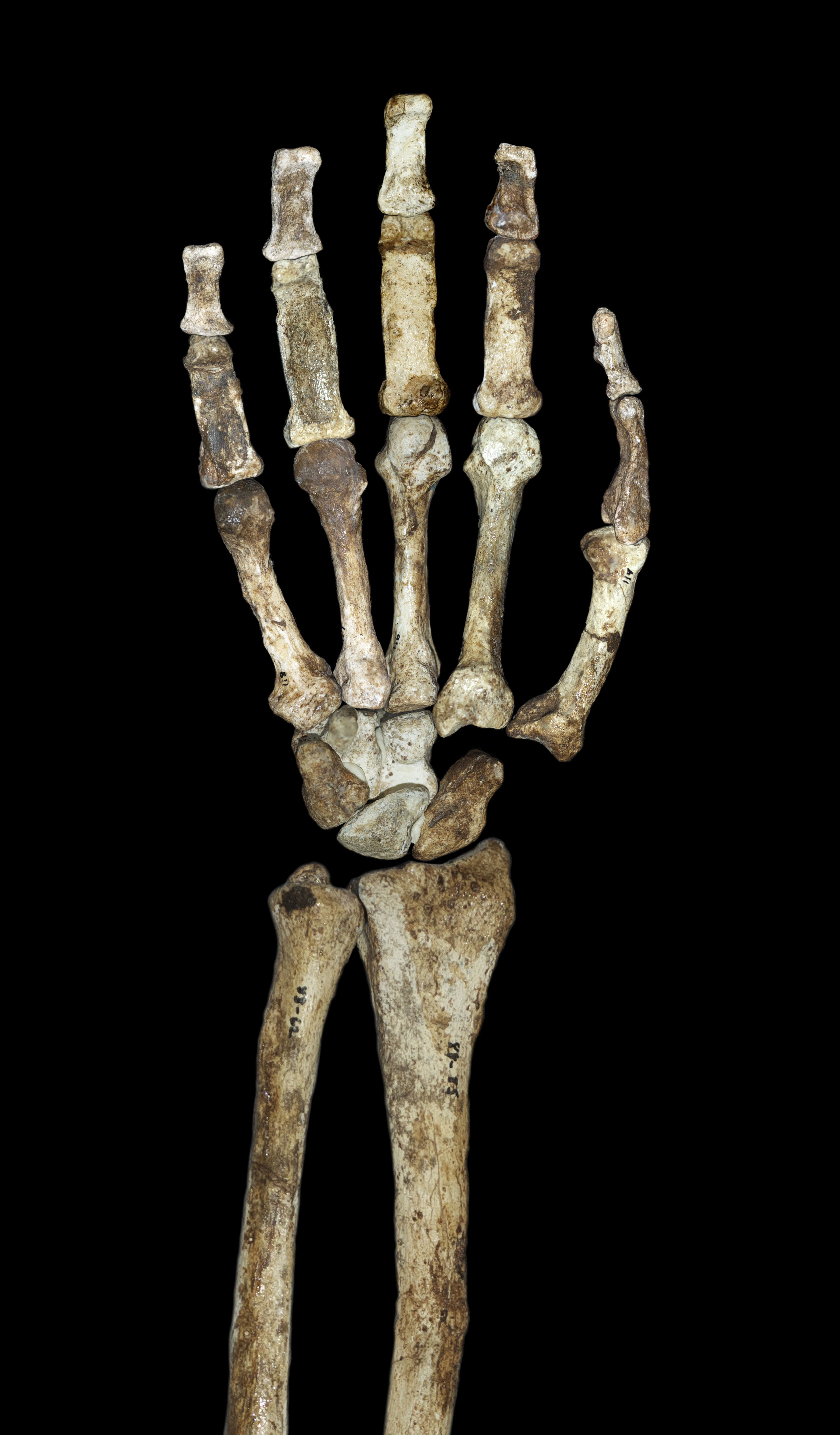
The hand of MH2, described by Kivell and colleagues as the most complete from any early hominin, displays a striking combination: curved finger bones associated with climbing alongside a long thumb and short fingers permitting a precision grip comparable to modern humans.3 The thumb-to-finger ratio was actually more favorable for precision gripping than in Homo habilis—the species whose name means "handy man."3, 13 Kivell and colleagues concluded this hand was a better candidate for the ancestral condition from which the later Homo hand evolved.3
Mosaic features of Australopithecus sediba2, 10, 11, 3
| Anatomical region | Australopithecine-like features | Homo-like features |
|---|---|---|
| Brain | Small volume (~420 cc) | Reorganized frontal lobes |
| Dentition | Small teeth relative to body size | Reduced tooth size compared to earlier australopiths |
| Hand | Curved fingers, strong flexors for climbing | Long thumb, precision grip capability |
| Pelvis | Overall small body size | Reshaped ilium suggesting human-like gait |
| Lower limb | Relatively long arms | Femoral biomechanics approaching Homo |
The debate over A. sediba and Homo ancestry
Berger and collaborators have consistently argued A. sediba is the best candidate for the ancestor of Homo, pointing to its unique combination of Homo-like brain, pelvis, hand, and dentition.2, 14 A 2012 cladistic analysis placed it as a sister taxon to Homo, closer than either A. africanus or A. afarensis.14
The most fundamental objection is chronological: at ~1.98 Ma, the Malapa specimens postdate the earliest Homo fossils (~2.33 Ma).15 A species cannot be ancestral to a lineage that already existed. Berger has responded that the known specimens need not be the earliest members of their species—older, undiscovered populations may have existed.14
Others question whether A. sediba is distinct from A. africanus at all, arguing the differences fall within expected variation for a single lineage over several hundred thousand years.9 A 2016 study further cautioned that MH1's juvenile status makes it unreliable for taxonomic assessment.16
Despite these objections, the Malapa fossils demonstrate conclusively that the australopithecine-to-Homo transition was mosaic, with different body systems evolving at different rates and in different sequences.14, 17
Australopithecus deyiremeda: coexistence in the Pliocene
Australopithecus deyiremeda was described in 2015 from jaw fragments at the Woranso-Mille site in Ethiopia's Afar region.18 The name combines the Afar words for "close" and "relative."18 Dated to 3.3–3.5 million years ago—contemporaneous with A. afarensis—its discovery provided the first compelling evidence that multiple hominin species coexisted within the same temporal and geographic range during the middle Pliocene.18
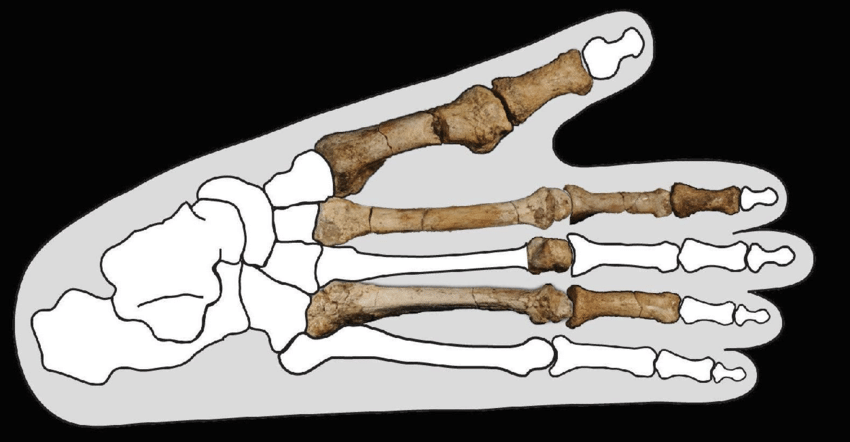
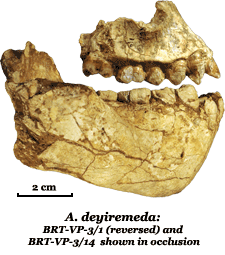
The holotype (BRT-VP-3/1) preserves teeth differing from A. afarensis in several respects: smaller anterior teeth, more primitive canine and premolar morphology, and a different enamel thickness pattern.18 Some researchers counter that these may represent extreme variants within a highly variable A. afarensis population.19
A major breakthrough came in late 2025, when new postcranial remains published in Nature included a partial foot with a divergent big toe suited to arboreal locomotion—unlike the flatter, more human-like foot of A. afarensis.20 Carbon isotope analysis showed A. deyiremeda relied primarily on C3 foods from trees and shrubs, contrasting with the mixed C3/C4 diet of A. afarensis.20 These findings demonstrated ecological niche partitioning, explaining how the two species coexisted without competitive exclusion.20
A diverse radiation, not a linear chain
These three species, combined with A. afarensis, A. africanus, and the robust Paranthropus lineage, reveal the late Pliocene and early Pleistocene as far more diverse than recognized even two decades ago. Between 3.5 and 2.0 million years ago, at least four and possibly six or more australopithecine species lived simultaneously in eastern and southern Africa.19, 21
Estimated cranial capacities of later australopithecines compared with related species1, 2, 22
The traditional model of a single australopithecine gradually transforming into Homo has given way to a scenario in which multiple lineages experimented with different strategies.21 Paranthropus developed massive chewing apparatus; the Homo lineage invested in larger brains and tools. The species described here explored their own combinations: A. garhi with megadont teeth and apparent tool use, A. sediba with a human-like hand and brain reorganization but small brain, and A. deyiremeda with an arboreal foot and specialized C3 diet.1, 2, 20
The recognition that Homo emerged from this diverse community rather than a single ancestor-descendant chain is one of the most significant conceptual shifts in paleoanthropology. The question is no longer whether multiple species coexisted but how they partitioned the landscape and which gave rise to Homo.21
Key specimens
Key specimens of later Australopithecus species1, 2, 3, 18
| Specimen | Species | Age | Location | Significance |
|---|---|---|---|---|
| BOU-VP-12/130 | A. garhi | 2.5 Ma | Bouri, Ethiopia | Holotype cranium; ~450 cc; associated with butchered bones |
| MH1 "Karabo" | A. sediba | 1.98 Ma | Malapa, South Africa | Juvenile holotype; ~420 cc; reorganized frontal lobes |
| MH2 | A. sediba | 1.98 Ma | Malapa, South Africa | Adult female; most complete early hominin hand; precision grip |
| BRT-VP-3/1 | A. deyiremeda | 3.4 Ma | Woranso-Mille, Ethiopia | Holotype maxilla; evidence of hominin coexistence in Pliocene |
Significance for human evolution
These species collectively demonstrate several principles central to modern understanding. First, evolutionary change was mosaic: brain reorganization may have preceded brain enlargement, pelvic changes preceded obstetric demands, and hand dexterity emerged before sustained stone tool production.10, 11, 3
Second, the boundary between Australopithecus and Homo is not a sharp line but a zone of overlap. A. sediba possesses features that, found in isolation, would be assigned to early Homo—yet the overall package remains more australopithecine.2, 14 This blurring is exactly what evolutionary theory predicts: species change through gradual accumulation, not instantaneous transformation.17
Third, the coexistence of multiple species in the same landscapes confirms that hominin diversity was the norm, not the exception.18, 20, 21 Today's situation—Homo sapiens as the sole surviving hominin—is the anomaly, having existed for fewer than 50,000 of the seven-million-year hominin record.23 The later australopithecines remind us the path to our species was neither straight nor solitary.
References
Australopithecus sediba hand demonstrates mosaic evolution of locomotor and manipulative abilities
2.6-Million-year-old stone tools and associated bones from OGS-6 and OGS-7, Gona, Afar, Ethiopia
Late Pliocene Homo and Oldowan tools from the Hadar Formation (Kada Hadar Member), Ethiopia
Australopithecus sediba and the emergence of Homo: Questionable evidence from the cranium of the juvenile holotype MH 1
Middle Pliocene hominin diversity: Australopithecus deyiremeda and Kenyanthropus platyops